Notes on Regeneration
Salamanders, especially the sub-group of salamanders called newts, can regenerate their legs.
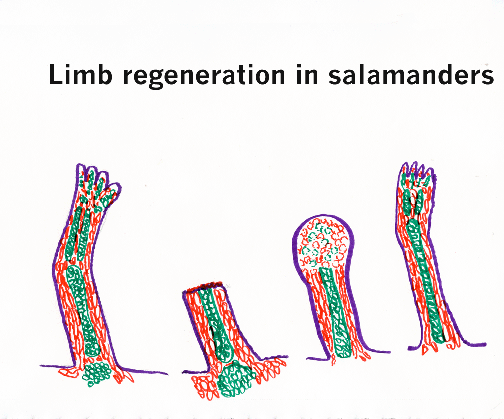
Their legs are not really simpler than mammal legs. So it is a good question why humans can't grow back arms and legs that somehow got cut off? Maybe researchers will discover some fundamental reason why it will forever be fundamentally impossible for mammals to regenerate legs and arms. Another possibility is that one of you in this class will discover a method to stimulate regrowth of arms and legs.
A specific question is whether differentiated cells switch from one cell type to another (for example, whether descendants of skeletal cells sometimes become muscle cells, etc.)
The ability of cells to switch from one cell type to another could possibly be the limiting variable, needed for limb regeneration to occur! However, radioactive labeling studies on salamander leg regeneration show that skeletal cells " dedifferentiate" in the " blastema" but then always redifferentiate back into skeletal cells; likewise, muscle cells seem to dedifferentiate, but when they redifferentiate they only become muscle; also skin cells become skin etc.
It seems to me (but I would welcome counter-arguments) that if all the skeletal cells of a regenerated leg are descended from cells that had been skeletal cells in the stump of the cut off leg, and ditto for muscle cells, skin cells, endothelial cells, etc. then wouldn't that mean that regeneration of limb structures amounts to a form of cell sorting? The descendants of cells that had been part of the femur, first dedifferentiate, then divide, and then redifferentiate as skeletal cells, but this redifferentiation is somehow in the geometric pattern of the radius and the ulna, and also the geometric patterns of wrist bones, finger bones etc. Likewise, for muscles, cells that had been part of biceps muscles would seemingly dedifferentiate, then divide to help form the blastema, and when they redifferentiate somehow form the geometric pattern of muscles in the lower arm.
Is this situation analogous to H. V. Wilson misinterpreting the reformation of dissociated sponges as being caused by cells switching from being one cell type to being a different cell type, according to their new locations? What do you think? You don't need to agree with me! Just understand the issue.
Serious medical journal articles have reported successful regeneration of the last joint of human fingers, as long as the cut stump isn't too tightly bandaged.
Another good question is why cuts and scratches heal so much faster in children than they do in adults. If you want a demonstration of this phenomenon, all you need to do is to take some children black-berry picking! You and they will get lots of scratches on your hands and arms, from the thorns. Three or four days later, your scratches will have begun the process of healing, and the kids' scratches will have healed. I speak from personal experience on this; the contrast is amazing; and you know for sure that you and the children got the scratches at the same time.
Does regeneration require removal of some inhibitory signal?
At least one researcher has proposed that skin produces an inhibitor of its own growth and regeneration. Analogous to the way in which auxin was first invented as a hypothetical substance, and then eventually isolated and identified by means of a bioassay, can you invent a bioassay to detect and compare amounts of s substance that inhibits skin cells from growing and dividing?
A British researcher invented the name "chalone" for this hypothetical self-inhibitor of skin growth. He pulled away the outer layers of mouse skin (epidermis), and measured increases in rates of growth and division of the basal layers of epidermis. Then he tested chemicals isolated from skin and fractionated, for example by chromatography and/or electrophoresis. He reported some positive data, indicating that he was getting close to the discovery of some one particular substance, used by skin to inhibit its own growth rates. But chalone still has not been proven to exist. Maybe a mixture of chemicals produce the effect; maybe the real mechanism works by destroying a self-stimulating substance.
Maybe the answer to regenerating legs and arms will depend on discovering what mechanism inhibits tissues from growing too much, until tissues are removed?
Can you invent a (potentially!) practical bioassay for such a hypothetical self-inhibitory substance, for salamander limb buds?
In terms of the existence and sensitivity of limb tissues to such a self-inhibitory substance (that is, IF it really existed, and IF reduction in amounts of such a self-inhibitory substance actually was how limb regeneration was caused and controlled) how would you explain the greater regeneration ability of salamander legs, as compared with human leg?
If such a substance really was the means of control of leg regeneration, suggest research that could lead to practical medical treatments that could cause regeneration of human arms and legs.
An excellent recent review paper on this topic:
Stocum, D.L., and Cameron, J.A. (2011). Looking proximally and distally: 100 years of limb regeneration and beyond. Developmental Dynamics 240(5), 943-968. PMID 21290477
Asexual Development by Budding, and Sorting Out of Dissociated Cells
What these phenomena have in common with each other is that bodies or parts of bodies can sometimes be formed in different ways than by embryonic development of an egg.*******************************************************************
Will NOT consider development of unfertilized oocytes, as occurs in production of male ants & bees, etc. and many other insects, and in some species of fish and lizards. Development of unfertilized eggs IS asexual, but is accomplished by gastrulation, neurulation, germ layers etc. almost exactly as in normal embryonic development. Sometimes results are haploid; but in many cases they become diploid by resorbing second polar body.
*******************************************************************
Development of buds of somatic tissues into whole new animals
Occurs in sponges, hydra, many other Cnideria, like corals, Planaria, and many invertebrate phyla that live as colonies, like Bryozoa
All of these can develop either from fertilized eggs, or from buds of tissues within which cells rearrange (and sometimes differentiate to form all the cells of their bodies)
Including colonial sea squirts. (note, however, that half or more species of sea squirts are NOT colonial, and develop only from fertilized eggs.
Budding of colonial sea squirts: Transfer of many differentiated cells, through blood vessels, and/or by active cell crawling.
New individuals produced by budding produce eggs and sperm, and also more asexual buds.
John Allen, who recently earned his PhD in this department, observed that the ends of arms of sea urchin plutei sometimes bud off to form new blastula stage embryos, which can themselves develop into plutei.
"Sorting out" of dissociated and randomly inter-mixed cells.
In some species of colonial sea squirts (and also sponges, hydra and other cnideria, and larval sea urchins)
As H.V.Wilson was the first to discover (in sponges) and later in corals, randomly mixed cells can rearrange to form normally-functioning individual sponges, etc.
*******************************************************************
Dissociated amphibian embryo cells sort out by germ layer, and by subdivision of germ layers, but can't form frog or salamander bodies, just tissues and some organs, in abnormal geometrical patterns.
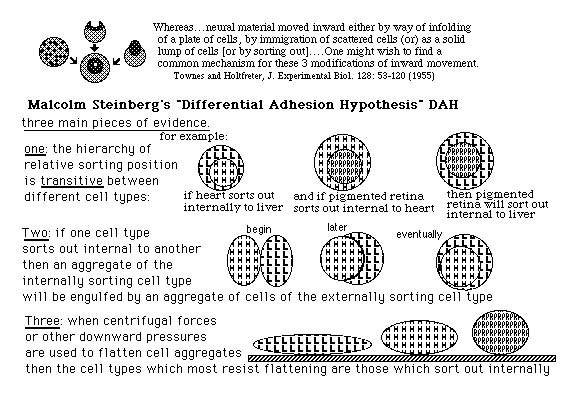
These phenomena were discovered and described in many research papers by Johannes Holtfreter. (especially a long paper by Townes & Holtfreter in J. Experimental Zoology about 1955, in which the results of all possible germ layer combinations are described)
In the 1950s, Moscona and Trinkaus (separately) proved that embryonic mouse and chicken tissues can be dissociated [nto random mixtures of cells, and then sort out by differentiated cell type (but canit re-form functional animals, but only tissues and organs)
This led to the discovery of selective cell-cell adhesion proteins, including N-CAM "Neural Cell Adhesion Molecule) and several kinds of Cadherins. ("Cadherin E, Cadherin P, many others)
*******************************************************************
Debates whether sorting out of dissociated cells is because of qualitative differences (in kind of adhesion molecules) or because of quantitative differences (in amount of adhesion)
Steinberg's "Differential Adhesion Hypothesis" << minimization of free energy of adhesion >>
evidence: transitive hierarchy of internal versus external position of sorting out
Alternative hypothetical explanations by Harris (1976) and Brodland (2005) "differential cell surface contraction hypothesis"
The non-specificity of any Bioassays that are based on inhibition of growth (or of any property) Lots of chemicals can inhibit, even if that inhibition is not their real function.
Imagine space aliens trying to identify the brakes on cars by damaging different parts, in search of some one part which will inhibit the car's forward movement.
What would they discover? That the fuel line is the brake? Or the carburetor / fuel injection?
Or the transmission? Or the inflation of the tires?
Every part needed for movement could be misinterpreted as being the brakes
****************************************************************************************
Should we think of regeneration as some kind of de-repression.
Alternatively, should we hypothesize that regeneration is a stimulation?
And if so, a stimulation by what? By damage? By the need for the missing organ?
Santayana said that those who do not learn history are condemned to repeat it.
I say that those who do not learn about this metaphysical puzzle (de-repression versus stimulation) are doomed to repeat it.
In relation to every example of organ regeneration.
****************************************************************************************
Another important question: What halts regeneration when it is finished?
An even more general question:
What halts normal embryonic development when it is finished?
Notice that this question is very important to medicine. For example, is it a useful way to think about cancer, as either failed repression or excess stimulation?
Hoping to find methods to cause regeneration (even of arms and legs!), will success come from stimulation?, or from removal of repression?
(Always remember Santayana's quote about those who don't remembering history, or those who never learned history in the first place.)
****************************************************************************************
Return to Wolffian Lens Regeneration, in search of general principles:
Drawings (will be done on the blackboard in class, and eventually posted on the web):
Wolffian Lens Regeneration sequence
1 before,
2 just after,
3 bulge and loss of pigment, partial
4 near completion
-----------
front view, noticing that regeneration is specifically from upper central part of iris
What happens if the dorsal-most iris is removed?
two lenses from sides?
one lens from ventral?
Can we make sense of this?
Gradation of some competitive capability?
Does that mean a competitive repression of some tendency to re-form lenses?
(or could it be a more positive, competition in which dorsal iris "wins",
unless removed, in which case the sides of the iris are de-repressed
Also, we should be curious why an organ doesn't regenerate until it is removed
Does an existing lens inhibit all parts of the iris from converting themselves into new lenses?
Apparently, yes. What could the mechanism be?
Maybe this mechanism is analogous to the hypothetical "Chalone" of skin (That Bullough hypothesized, and applied the bioassay method to try to identify, (unsuccessfully)?
Questions for class discussion:
A) What are some alternative reasons why chalone wasn't isolated?
1) Because it doesn't really exist?
2) ?
3) ?
B) Can we invent an equivalent to chalone as an explanation for Wolffian Lens Regeneration?
C) Would that include competition between dorsal iris versus lateral iris?
1) Maybe...
2) But not unless....
3) But only if....
4) Something else, that we haven't thought of yet
D) Would our mechanism (or some alternative mechanisms?) account for gradually bringing regeneration to a halt, when it is finished.
E) Invent possible bioassays that could identify the equivalent to chalone for lens regeneration?
F) ....For Regeneration of Arms and Legs?