Contact Inhibition of Cell Locomotion: Nov 13, 2019
The following are good examples of developing quantitative, statistical criteria and measures for phenomena. (By Michael Abercrombie and Joan Heaysman)1) Speeds of fibroblast locomotion are reduced in cells that are touching other cells (For example, those touching six other cells move only half as fast as those touching zero other cells).
2) When cells crawl outward from two nearby tissue fragments the number of cells per area between the two tissue fragments is less than twice as many cells per area as compared with locations at the same distance outward from only one tissue fragment.
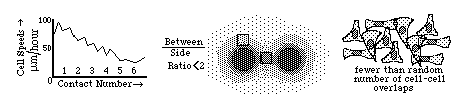
3) Count observed overlaps between cells, and compare with calculations of numbers of cell-cell overlaps that would/should have occurred if cell locations were random (There were only about half as many actual overlaps, as there would have been if cell locations were random). A result is formation of cell "monolayers" one cell thick, within which locomotion almost stops.
4) Collaboration with Jack Ambrose, using "Interference Microscopy" to compare cell surface movements at locations where one cell crawls into contact with another cell. "Ruffled Membranes" are locally inhibited, but often with less inhibition when cancer cells collide. Membrane ruffling was previously hypothesized to be peristaltic waves in the cell surface; but then hypothesized to be the result of concentrated local reassembly of cytoplasmic actin along spreading edges of crawling cells.
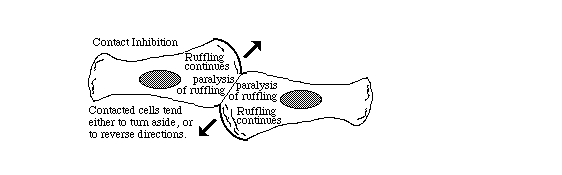
Either way, cells behave as if their motors were inhibited or re-directed away from locations where one cell touches another. This has been called "contact paralysis". Unfortunately, nobody yet understands the "motors" by which tissue cells crawl. It includes * Actin assembly, ** retrograde transport of attached particles, *** membrane flow, and **** traction force measured by wrinkling of rubber and gel substrata.
Inhibition of these processes is concentrated at sites of cell-cell contact, and does not expand to nearby areas (showing that the cause can't be diffusion of an inhibitor or an electrical depolarization.)
Other researchers compared rates of mitotic cell cycling in crowded cultures as compared to cultures in which these is less cell-cell contact. They discovered what they named "contact inhibition" of cell growth; and also observed that this inhibition was reduced in cancer cells. This stimulated much excitement.
Soon, when scientists wrote or talked about "contact inhibition", what they usually were referring to was the inhibition of growth.Next it became recognized that crowding of cell cultures inhibited growth , whereas growth and mitoses were increased every time cultures were fed with fresh medium. The inhibition of growth may not result from contact (despite the name). Soon after that, many decided that "contact inhibition had been disproven".
--------------------------------------------------------------
Please read this paper:
http://www.pnas.org/content/115/42/10678.long
Li and Wang (2018) "Coordination of cell migration mediated by site-dependent cell-cell contact" PNAS vol. 115, pages 10,678 - 10, 683.
ABSTRACT: Contact inhibition of locomotion (CIL), the repulsive response of cells upon cell-cell contact, has been the predominant paradigm for contact-mediated responses. However, it is difficult for CIL alone to account for the complex behavior of cells within a multicellular environment, where cells often migrate in cohorts such as sheets, clusters, and streams. Although cell-cell adhesion and mechanical interactions play a role, how individual cells coordinate their migration within a multicellular environment remains unclear. Using micropatterned substrates to guide cell migration and manipulate cell-cell contact, we show that contacts between different regions of cells elicit different responses. Repulsive responses were limited to interaction with the head of a migrating cell, while contact with the tail of a neighboring cell promoted migration toward the tail. The latter behavior, termed contact following of locomotion (CFL), required the Wnt signaling pathway. Inhibition of the Wnt pathway disrupted not only CFL but also collective migration of epithelial cells, without affecting the migration of individual cells. In contrast, inhibition of myosin II with blebbistatin disrupted the migration of both individual epithelial cells and collectives. We propose that CFL, in conjunction with CIL, plays a major role in guiding and coordinating cell migration within a multicellular environment.