Embryology Biology 441 Spring 2012 Albert Harris
Monday, March 26, 2012
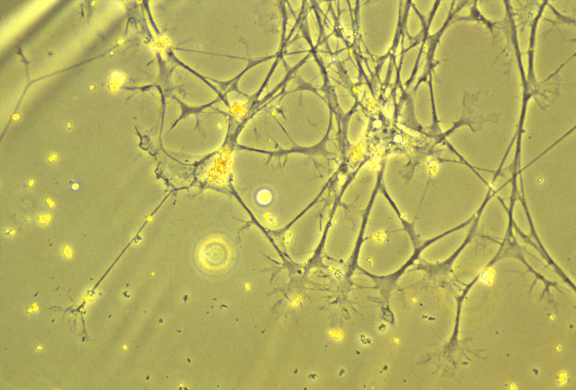
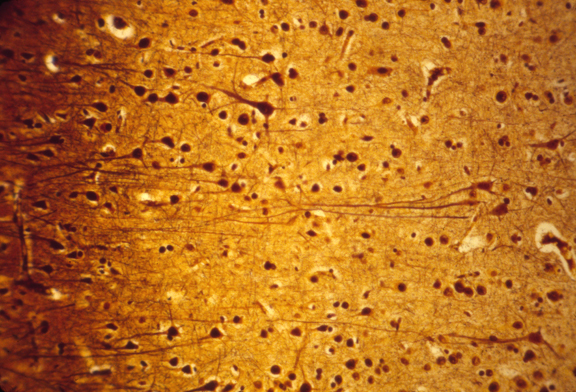

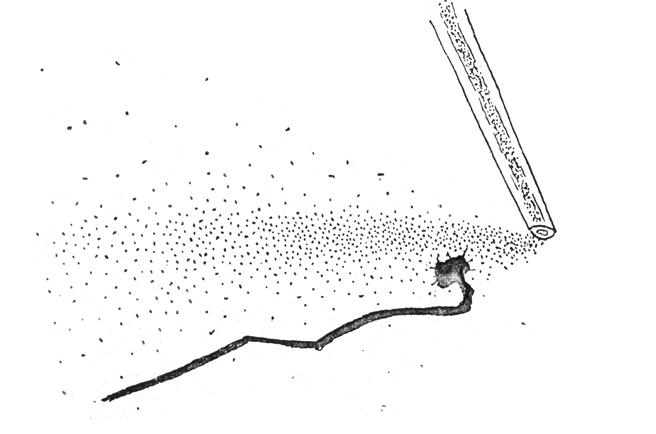
Nerve cell ("neuron")
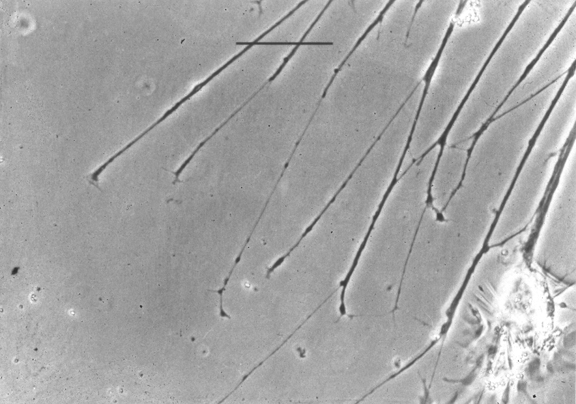
Nerve growth cone: amoeboid tip of future axons and dendrites.
Axon paths are caused by active locomotion of growth cones.
The thin strand of cytoplasm they leave behind becomes an axon.
Motor nerves develop from neural tube ectoderm
Sensory nerves (of the skin, etc. ) develop from neural crest ectoderm
Autonomic nerves (that control intestines, heart rate, blood pressure,)
Also develop from neural crest
(The post-ganglionic autonomic nerves are from neural crest)
(The pre-ganglionic autonomic nerves are from neural tube)
Please notice that autonomic nerves are a special kind of motor nerve.
_____________________________________________________________________
1) Please imagine a science fiction story in which the internal wiring of very powerful computers is done by amoeboid locomotion.
2) These "amoebae" are the tip ends ("growth cones") of very long thin strands of cytoplasm.
3) The "amoebae" are at least partly guided by combinations of concentration gradients of certain proteins on the surfaces of target cells.
4) These gradients are not diffusion gradients, but it is possible that diffusion gradients cause their formation.
5) Quantitative gradients can be caused by many kinds of sequential equilibration between neighbors.
6) Growth cones reach their targets, and become synapses.
7) The strands of cytoplasm become nerve axons.
[Axons can conduct action potentials equally well in both directions; but synapses can't.]
8) The proper wiring of these science fiction computers ("brains') result mostly from the
two (or more) ends of each nerve fiber forming synapses with the correct other cells.
e.g. Whatever nerve cell forms a synapse with cell Q at one end should connect with cell q at the other end. (And the same happens for connecting to R at one end and to r at the other, s to S, t to T etc. etc.
9) This can be accomplished in at least 4 fundamentally different ways, 3 of which people often ignore:
-
One) Each nerve cell can somehow becomes special in a way that causes its ends to find and connect to Q at one end, and connect to q at the other end.
Two ) Nerve cells can connect somewhat randomly, and whatever cell connects to Q is caused to change in some way that a signal propagates down the length of the fiber and causes the other end to seek out and connect to q.
Three) The reverse of 2; whoever connects to q is changed to cause it to seek out Q.
Four) Nerves experiment repeatedly at both ends, as a human might test a circuit with a voltmeter. One end tries connections with a, b, c, q etc. and the other end of the nerve tries connections with Z, Y, X, W, Q etc. until the two ends happen to match, such as a nerve simultaneously connecting to m and to M. At that point, synapses would become more permanent, and stop experimenting.
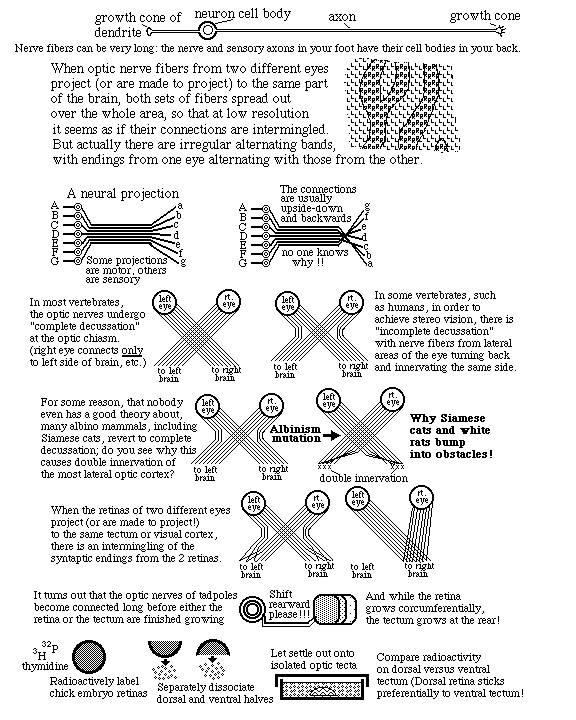
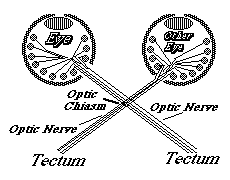
11) Ganglion cells extend axons toward the "blind spot", where the optic nerve leaves the back of the eye, and where there are no rods & cones, and no vision. Nobody knows (or asks) what guidance mechanism steers the fibers toward the blind spot. But there must be some mechanism.
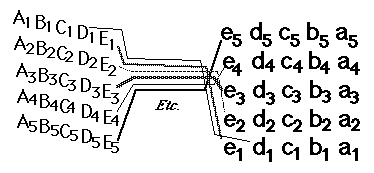
12) From the blind spot, the optic nerve growth cones crawl together, side by side, under the brain, passing perpendicularly among nerve fibers coming from the other eye, and up the other side of the brain, eventually spreading out across the upper surface of the midbrain ("the optic tectum"), and forming a neural projection (map-like geometric pattern of connection).
13) This "retino-tectal projection " is one of dozens or maybe hundreds of map-like patterns of connection in the human nervous system. We can't be sure they are all caused by the same basic mechanism; but maybe they are, except with differences in adhesion proteins.
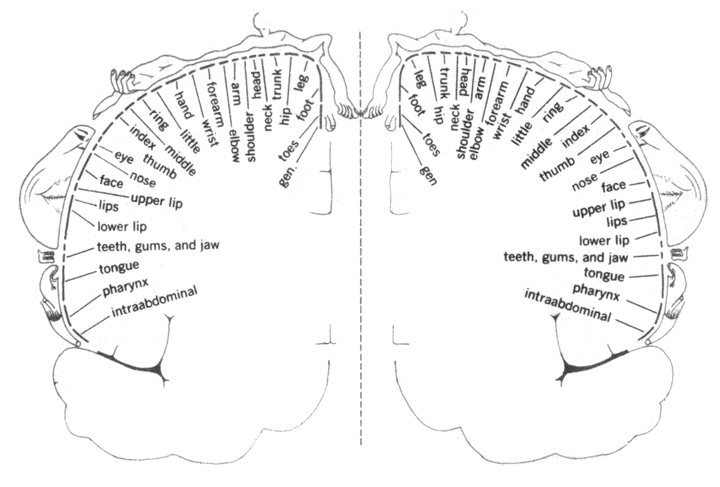
14) Two other examples of neural projections, one of sensory nerves and the other of motor nerves are illustrated by a pair of diagrams in many Psychology textbooks, that shows the whole human body spread out across parts of the cerebrum. Where the diagrams of the face are drawn means those are the parts of the brain that receive nerve impulses from the face (probably relayed via several nerves).
15) Some history of embryological research on causation of the retino-tectal projection. (attempts to discover what mechanisms guide ganglion cell axons to their correct part of the midbrain)
I) Salamander eyes were surgically removed, rotated 180 degrees and put back in. (1930s & later)
After a few months, the salamander regains vision in the rotated eye & sees everything backward.
The same experiment was later done with frogs, as diagramed on page 491 of the textbook.Mitoses by pigmented retina cells regenerate the neural retina, and these new nerves connect to brain!
II) * Chick embryo eyes were labeled with radioactive isotopes, cut into dorsal and ventral halves
dissociated into individual cells.
** Chick embryo midbrains were dissected out and pinned to the bottom of Petri dishes in saline.
*** Dissociated retinal cells from dorsal retinas were pipetted onto the midbrains.
**** After a set time, not-yet-attached retinal cells were washed off of the midbrain.
***** Measurements of radioactivity showed that more dorsal retina cells stuck to lateral midbrain.
And more ventral retinal cells stuck to more medial parts of the midbrain.
i.e. Retinal cells stick about twice as well to the general part of the brain that axons from there would have innervated.
This was taken as evidence of selective adhesiveness. (And as evidence that neural projections are caused by quantitatively selective adhesiveness) (Prof. Steve Roth, Dept of Biology, U of Penn, was the leader of this research) (They never identified what proteins caused this adhesion)
III) Membranes were isolated from anterior parts of the optic tectum, and other membranes from posterior parts of the optic tectum. These isolated molecules were, in effect, painted onto glass surfaces, in alternating stripes (Fig. 12.26) and retinal cells were put in tissue culture next to them. Nerve cells from posterior retina extended axons much farther onto stripes of anterior brain stuff.
i.e. Nerve axons crawl much farther on molecules from the area of brain they would have innervated.
The Chemoaffinity theory of Roger Sperry was confirmed by results like this. (Sperry won a Nobel Prize, partly for this & partly for human "split brain" surgery for epilepsy.) The affinity could be either selective adhesiveness ("haptotaxis ") or more like chemotaxis .
Either of which could be turning of cell locomotion toward increasing chemical concentration, or could be decreasing/increasing of turning, or decreasing vs. increasing of retraction .
Scientists tend to think of chemotaxis as ONE mechanism, But bacterial chemotaxis is by increase in random turning when attractant concentration decreases.
Leucocyte & Dictyostelium chemotaxis is by turning of locomotion toward side where higher concentration of attractant.
Fibroblast haptotaxis is caused by a tug of war between opposite sides of cells.
(& I got hired at UNC for doing experiments that prove that cause. (Exp. Cell Res. 1973) An alternative idea was that cells were pulled by maximization of adhesions. That theory is still believed by most to be the cause of cell-type-specific cell sorting. Although I have published papers arguing that active contraction is the driving force)
I wish I had been smart enough to realize that induced detachment is a possible mechanism. By ephrin protein (on retinal cells) and ephrin receptor protein (on tectal cells)
The more ephrin a cell has on its surface, the less ephrin receptor will be needed to induce retraction.
The less ephrin a nerve cell has on its surface, the higher the concentration of ephrin receptor will be needed to induce detachment and retraction of nerve growth cones. Figure 12.27
As an analogy, consider people running across really hot beach sand, toward the water. If people differ in sensitivity to pain, who will stop running closest to the ocean? If there is a gradient of temperature of the sand? Which direction would this sand temperature gradient need to increase?
Our textbook refers to induced de-adhesion as "repulsion" (which seems misleading to me)
Long range effects of diffusible chemicals have also been discovered ("repulsion" & "attraction")
Plus there are perpendicular gradients of ephrin A and ephrin B, which each have their own specific receptors
******************************************************************************
Questions for class discussion:
#1) What do you think actually was happening in Steve Roth's experiments?#2) What was really happening in the experiments with alternating stripes of isolated membranes from anterior versus posterior parts of the midbrain?
#3) The retino-tectal connection pattern is upside-down and backwards. Ant. retina to posterior tectum, Dorsal retina to ventral tectum, etc. etc. I have always suspected this was some kind of clue about the causal mechanism (like Chargaff's rules) But I never could figure out what! Look at figure 12.27 Can you figure out a reason why upside-down and front-to-back make sense in terms of ephrins? (and of induction of retraction, or "repulsion".)
#4) In haptotaxis studies of cells crawling on adhesion gradients, all differentiated cells except macrophages accumulate preferentially on whichever alternative surface material is more wettable ; but macrophages accumulate preferentially on whichever material is least wettable
Can you figure out possible reasons that could explain this?
#5) Osteoclasts develop from macrophages : should somebody test if they accumulate preferentially on more hydrophobic plastics? (And test if osteocytes do the opposite?)
How could the result help cure & avoid osteoporosis?
Retino-Tectal Projection
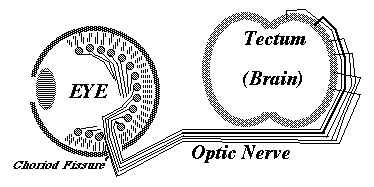

Embryonic eyeball, with retina surrounding it. The groove is the location of the optic nerve.
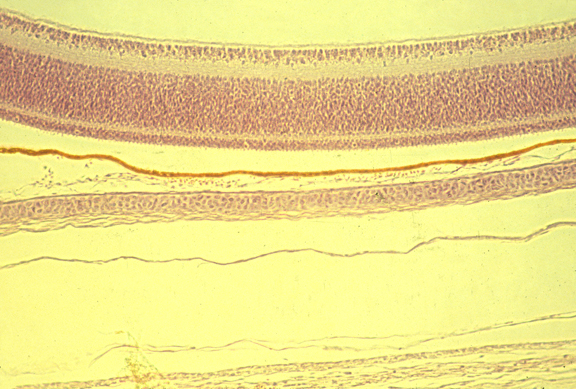
Cross-section of developing retina
Cross-section of optic nerve