Monday, March 5, 2018
Cell Sorting and Discovery of Selective Adhesion Proteins
Sorting out by randomly remixed differentiated cells of sponges, coral, Hydra, and embryos of echinoderms, amphibians, birds and mammals.H.V. Wilson (1907). Henry Van Peters Wilson of the UNC Dept. of Biology discovered sorting out by dissociated and randomly re-mixed cells of living sponges. In 1911 Wilson reported the same phenomenon in cells dissociated from soft corals.
There is a good painting (oil portrait) of Wilson across the hall from this lecture room.

This photograph shows the painting of Wilson, with Nelson Hairston and Calhoun Bond in the foreground.
Dr. Hairston was Wilson's last graduate student, in the late 1930s, and became a very famous ecologist.
Cal Bond did his Ph.D. research in my laboratory.

Oil painting of Albert Harris by his father Kenneth Harris,
painted about the time the future Professor was first
getting interested in Biology.
NOTE: Wilson did not believe that cells rearrange according to cell type; that idea was later proposed by Julian Huxley, based on his own observations of dissociated sponge cells.
Wilson believed that dissociated sponge cells switch from one differentiated state to another, depending on cell locations.
Does position control differentiation? (As Wilson argued against) (Cells at the surface of aggregates switch cell type to become skin, etc.) Or is re-positioning of cells controlled by which differentiated cell type they are? (Skin cells move from the interior back to the surface, re-forming skin.)
Later Wilson published papers arguing that sponges reform by differentiation of previously undifferentiated "archeocytes" (analogous to stem cells). (This is analogous to Columbus arguing that he had reached China, not discovered new continents.)
Wilson tried mixing dissociated cells from sponges of different species, trying to make super-sponges that might be economically useful. It didn't work. Cells of different sponge species separated from each other.
This is in contrast to what happens if you dissociate and mix cells from salamanders and frogs, or from birds and mammals.
Dissociated salamander and frog tissues adhere together, and then sort out into layers according to whether they are neural versus somatic ectoderm, or mesoderm versus endoderm, etc. This was discovered by Johannes Holtfreter in the 1930s, 40s, 50s & 60s.
Holtfreter interpreted sorting out as resulting from differences in "Tissue Affinity", which led other scientists to discover cadherins, N-cam, etc., which are selective cell-cell adhesion proteins.
The classic paper is Townes and Holtfreter (1955), Directed Movements and Selective Adhesion of Embryonic Amphibian Cells, Journal of Experimental Zoology 128, pp. 53-120. If you have a long weekend sometime, I can recommend it.
Embryonic bird heart and liver cells can be dissociated and randomly re-mixed with heart and liver cells from embryonic mice, with the result that heart cells sort out to internal clumps, surrounded by layers of liver cells. Heart moves to be with heart, despite being mouse and chick, etc. This was demonstrated by Aaron Moscona. (1950s & 60s)
J. P. Trinkaus (1950s, 60s & later) used radioactive labeling of DNA to prove that dissociated cells were re-arranging, not re-differentiating. Using pigmented retina cells mixed with other cell types, Trinkaus confirmed the same conclusion, and produced time lapse movies in which sorting out of pigment cells was directly observable. But Trinkaus rejected the idea that normal cell rearrangements (gastrulation, neurulation etc.) are caused by the same mechanisms as sorting out of dissociated cells. (as was argued by several of his graduate students, who had gone to his laboratory hoping to use sorting as a pathway toward explaining gastrulation, neurulation, somite formation, etc. Trinkaus' reasoning was that retina sorts out from limb bud, even though these tissues never normally contact each other (& likewise many other combinations of cells sort out, despite never normally being in contact! So why would any normal mechanism produce results that never normally occur? His students were not convinced.)
Malcolm Steinberg (1964, and later) proposed a "thermodynamic" theory that sorting out of dissociated chick, mouse and amphibian cells is caused by quantitative differences in amount of cell-cell adhesiveness (amount rather than kind). The Differential Adhesion Hypothesis or DAH.
This theory had the great advantage of making testable predictions. Relative interior-exterior positions of sorting out should form a "transitive hierarchy."
If cell type A sorts out to interior positions relative to cell type B, and if B sorts out interior to cell type C, then (for every combination) A will sort out interior to C.
Steinberg tried more than forty different pairwise combinations of two differentiated cell types per experiment, and their relative positions, interior versus exterior, was always transitive. If cell type A is more adhesive than B, and B more adhesive than C, then A will be more adhesive than C. Therefore the variable that causes position in sorting must be amount of adhesiveness.
A critic argued that transitivity is predicted by any quantitative variable. If A were more strongly contractile than B and B were more contractile than C, then A would be more contractile than C. Differences in amount of anything makes the same prediction of transitivity.
Another set of testable predictions had to do with resistance of cell aggregates to flattening by centrifugal (or other) forces. The key ideas were that adhesion is close-range atraction, and that the rounding up of multicellular masses (equivalent to "surface tension") was driven by cell-cell adhesive forces. Therefore, one ought to be able to measure the force of cell-cell adhesion based on resistance of cell aggregates to flattening by centrifugal forces some give strength.
Aggregates of heart cells, liver cells, etc. were centrifuged, and the amounts of flattening were measured. Liver aggregates flattened more than aggregates of heart cells, and so on: The same transitive hierarchy resulted. Therefore Steinberg believed he had proven that cell-cell adhesiveness must create the driving force that causes cell aggregates to round up.
The same critic mentioned above suggested that the driving force that causes re-aggregating cells to round up might be acto-myosin based contraction, the same forces by which liver cells, heart cells etc. crawl. In other words, maybe the reason that cell aggregates behave as if they have a contractile surface layer is because they DO have a contractile surface layer. To account for observed cell behaviors, cell contractility would need to be inhibited on those parts of cell surfaces which touch other cells, and that less inhibition is produced by contact with cells of a different differentiated cell type as compared with the effect of contact with the same cell type.
Many scientists doubt that sorting out and rounding up have anything much to do with normal cell behaviors. On the other hand, cancerous cells were discovered to sort out to the exterior position relative to the closest non-cancerous cells. Is it not important to find out whether cancer cells are less adhesive, or less contractile, than normal cells?
Perhaps motivated by the criticisms mentioned above, Steinberg made a sabbatical year visit to Takeichi's laboratory in Japan and studied mouse cancerous cells into which different numbers of genes for a cadherin had been inserted (1987). The cells with more copies of a cadherin gene sorted out to interior positions relative to cells with fewer copies of that gene. This is what the theory had predicted. They didn't test whether surface contractilities changed.
Steinberg claimed that his theory was "based on thermodynamic principles". The following are quotes from Scott Gilbert's mostly-excellent Embryology textbook: [link to this section in Gilbert's book
"What forces direct cell movement during morphogenesis? In 1964, Malcolm Steinberg proposed the differential adhesion hypothesis, a model that explained patterns of cell sorting based on thermodynamic principles.
"Cells interact so as to form an aggregate with the smallest interfacial free energy. In other words, the cells rearrange themselves into the most thermodynamically stable pattern.
In 1996, Foty and his colleagues in Steinberg's laboratory demonstrated that this was indeed the case: the cell types that had greater surface cohesion sorted within those cells that had less surface tension."
In my opinion, this is a misunderstanding of thermodynamics. The business about "rearranging into the most stable pattern" would only be (approximately) true if there are no additional sources of energy, like active contraction of acto-myosin. Steinberg defended his theory by vociferous claims that his critics were hopelessly ignorant of the basics of thermodynamics. The critic mentioned a couple times above happened to have suffered through a whole year of thermodynamics in college, and even understood a little of it.
C-P Heisenberg used an atomic force "microscope" to compare strengths of cell-cell adhesion versus forces of cell contraction in cells from fish embryos. The results were not what the DAH predicted.
M. L. Manning, Foty and Steinberg published a paper about "Co-Action" of cell contraction and cell adhesion, in the sense that both are partly true.
This paper has lots of equations which are not meant to be understood, but to continue the "emperor's (invisible) clothes" strategy of deflecting criticism.
Manning, M.L., R.A. Foty, M.S. Steinberg, and E.-M. Schoetz (2010). Coaction of intercellular adhesion and cortical tension specified tissue surface tension. PNAS 107, 12517-12522.
Can adhesion exert forces? Yes. But accumulation of cells preferentially on a substrate doesn't necessarily mean they were pulled there by adhesion.

Palladium metal was deposited in a grid pattern on both glass (left half of field)
and cellulose acetate (right half of field). Cells have accumulated on the palladium in
preference to cellulose acetate, but on glass in preference to palladium.
From Harris, A.K. (1973), Behavior of cultured cells on substrata of variable
adhesiveness. Experimental Cell Research 17, 285-297.
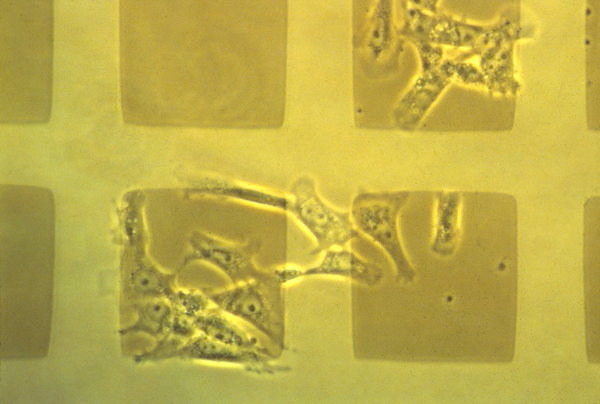
The cells here are clearly accumulating preferentially on the more sticky places,
but if you make movies, you see that the cells aren't being pulled by adhesion forces,
but are crawling around and then letting go on the less sticky areas: